“这项工作主要研究了摄食调控领域最受关注、被研究得最为清楚的一群神经元——下丘脑 AgRP 神经元。该研究最主要的内容是颠覆了领域内的固有认知:即 AgRP 神经元对于维持摄食行为和体重并非不可或缺。”对于自己担任共同通讯作者的一篇论文,中国科学与技术大学教授占成表示。

图 | 占成(来源:占成)
据介绍,针对 AgRP 神经元的研究热度已经持续数十年,近年来仍然不时有相关论文发表于顶刊。这主要是因为其对摄食、体重及能量代谢有非常显著的调控作用。
20 世纪末期,学界通过分析肥胖小鼠的基因突变发现[1,2],发现携带褐鼠被毛颜色基因的某些等位基因的小鼠呈现出黄毛、肥胖和体长增加等特征,且这些特征主要是由编码正常褐鼠蛋白(Agouti)的嵌合转录本的普遍表达所引发。
随后,人们从小鼠和人类中分离出一种基因,编码了一个在大小和基因组结构上与 Agouti 几乎相同的蛋白质:Agouti-related protein(Agrp)[3]。
AgRP 主要表达在大脑、肾上腺,以及垂体中。ob/ob 肥胖小鼠 AgRP 表达量明显高于正常小鼠,在小鼠中过表达 AgRP 基因也会导致肥胖。这说明 AgRP 的表达水平与肥胖密切相关。
随后,该领域内的研究逐渐聚焦于大脑中的 AgRP 神经元。AgRP 神经元只分布在下丘脑弓状核,它可以共表达 AgRP 和 Neuropeptide Y(NPY)[4],并使用 γ-aminobutyric acid(GABA)作为神经递质[5]。
AgRP 和 NPY 都有非常强的促食欲效果[6-8],向脑室直接注射这两种神经肽都能显著促进摄食。出人意料的是,无论是 NPY-/- 敲除小鼠还是 AgRP-/- 敲除小鼠,或者是 NPY-/-:AgRP-/- 双敲除小鼠,它们的摄食、体重和生长发育都与正常小鼠没有明显差异[8,9]。
对此的一种解释是:小鼠发育过程中产生了某种代偿机制,弥补了 AgRP 和 NPY 敲除对摄食体重的影响。
为了避免发育代偿效应,2005 年学界通过在成年小鼠 AgRP 神经元上,选择性地表达细菌白喉毒素的人类受体并注射白喉毒素,结果发现几天之内 AgRP 神经元就被杀死,进而导致小鼠停止摄食并迅速减重,甚至饥饿至死[10,11]。
这些结果让人们相信:对于维持成年动物的摄食、体重和生长,AgRP 神经元是不可或缺的。相关论文也被广泛引用和复述,并成为领域内的重要共识之一,也写入教科书[12]。
2011 年,人们利用当时最新发展出来的光遗传学和化学遗传学手段,对 AgRP 神经元进行特异性的激活,结果发现这可以快速、显著地促进小鼠的摄食行为,甚至会引起小鼠暴食[13,14]。
2020 年,人们发现长期增强 AgRP 神经元活性则会导致动物超级肥胖[15]。这些结果进一步确定了 AgRP 神经元在摄食和体重调控中的核心地位。
当时人们一度认为,AgRP 神经元活动的增强,会促进摄食从而导致肥胖;而 AgRP 神经元活动的缺失,将让动物丧失食欲以至于停止摄食。
2015 年,学界出乎意料地发现,尽管饥饿刺激可以导致 AgRP 神经元活动增加,但在开始进食和进食过程中,AgRP 神经元活动并没有增加,反而出现下降[16-18]。
其中一种解释是:虽然 AgRP 神经元可能会编码饥饿感,但其神经元活动的增加,对于摄食行为的产生和维持并不是必要的。
值得注意的是,2020 年人们通过在 AgRP 神经元上过表达内向整流钾通道蛋白 Kir2.1,实现了 AgRP 神经元活性的长期抑制,而这并不会影响成年小鼠的体重和摄食[15]。这些结果的产生,也对 AgRP 神经元活动在维持摄食和体重中的必要性提出了质疑。
一直以来,占成课题组对于大脑调控摄食的神经机制非常感兴趣。2019 年,该团队曾开展一项关于脑干孤束核一群 NPY 神经元对于促进摄食的研究,他们发现当 NPY-Cre 小鼠的下丘脑 NPY 神经元遭到毁坏,并不会对动物摄食和体重起到作用。
鉴于上述结果与领域内的共识互相矛盾,他们怀疑可能是实验出了问题、或者是 NPY-Cre 小鼠不够特异,因此应该用 AgRP 特异的 Cre 小鼠进行实验。
2021 年,在“中国神经调控代谢”学术会议上,占成在与美国德克萨斯大学教授童青春交流的过程中,非常惊奇地发现他们两个实验室的结论很相似。
与领域内其他几位老师交流之后,大家一致认为这是一个非常重要的问题,值得仔细研究。
于是,占成和童青春两家实验室决定分头行动,用不同的方法损毁 AgRP 神经元,并仔细地确认损毁效果,力求实验结论的可靠性。
占成说:“在此过程中,由于我本人经历了工作变动,在中科大的实验室刚刚建立之时又逢疫情,使得研究工作一度进展缓慢。”
好在两个实验室时刻保持沟通,不定期进行线上会议交流。最终在大家的共同努力下,两支课题组分别确认了 AgRP 神经元损毁的特异性和有效性,并得出了一致的结论。
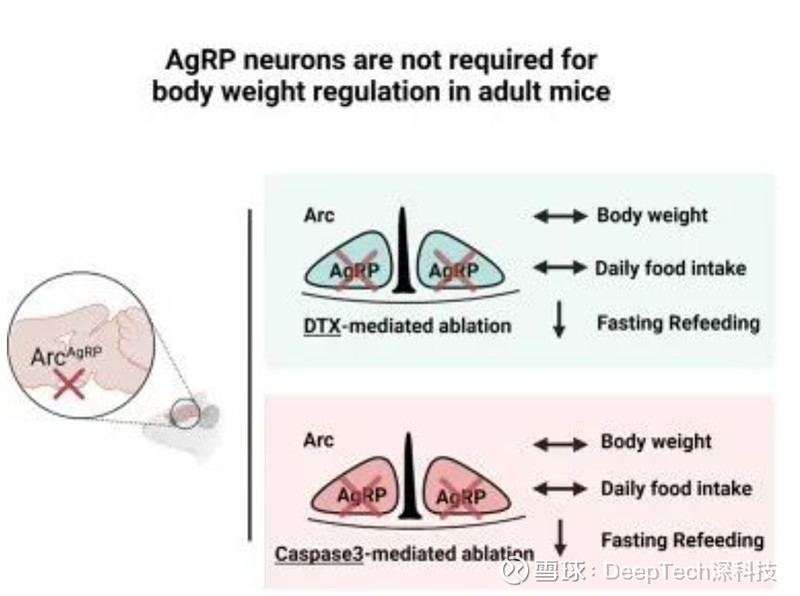
期间,童青春实验室通过在成年小鼠 AgRP 神经元上选择性地表达细菌白喉毒素的人类受体,并向脑室注射低剂量的白喉毒素,结果发现尽管这足以杀死 AgRP 神经元,却并不影响小鼠的体重和基础摄食水平。
而注射高剂量的白喉毒素,不仅能引起白喉毒素受体小鼠体重和摄食的下降,也会导致 WT 小鼠体重和摄食的下降甚至导致死亡。
为了避免白喉毒素的非特异毒性,占成实验室在 AgRP 神经元上选择性表达 Caspase-3 引起神经元凋亡。同样地,利用该方法杀死 AgRP 神经元,并不会降低成年小鼠的体重和摄食。由此可见,AgRP 神经元在维持实验室条件下的基础摄食和体重中并非是不可或缺的。随后,他们开始撰写论文并投稿。
(来源:Cell Reports)
占成表示:“论文投稿过程中,我们先是尝试了领域内多个知名期刊。有意思的是,我们的工作似乎更容易被知名度高的期刊送出去审稿,知名度较低的期刊则往往直接拒绝送审。”
他继续说道:“部分原因可能是我们的工作主要挑战了固有认知,即使是颠覆领域共识,但也看起来不像是一个新发现。也许只有对领域发展历史和现状有更好了解的审稿人才能更好地认可我们的工作,毕竟科学的进步不仅是探索新的知识,不断修正传统的认知也非常重要。”
最终,相关论文收录于Cell Reports,论文题目为《AgRP 神经元在成年小鼠体重维持中并非必不可少》(AgRP neurons are not indispensable for body weight maintenance in adult mice),柴静和陈静是共同一作,童青春和占成担任共同通讯作者。

图 | 相关论文(来源:Cell Reports)
总之,该研究颠覆了领域内对 AgRP 神经元的固有认知,将促使人们对 AgRP 神经元及其摄食行为的神经调控机制开展更加深入的研究。
本次成果在颠覆 AgRP 神经元必要性的同时,也说明摄食行为具有强“鲁棒性”,大脑调控摄食的机制很可能存在“冗余性”,即大脑内应该存在多个介导摄食的神经元。
事实上,过去几年人们已经发现多个具备促摄食功能的核团和神经元亚群。因此,深入了解不同神经元如何协调控制摄食,将有助于全面理解进食调控的神经机制,并为控制饮食和治疗摄食障碍提供新靶点和新思路。
下一步,占成等人将围绕摄食行为的“鲁棒性”和调控机制的“冗余性”开展研究。
一方面是要确定不同促食欲神经元之间地相互关联,另一方面则要寻找新的摄食调控靶点。
占成表示:“我们实验室的研究涉及神经、免疫、以及生理,主要开展神经-生理-免疫的交叉型研究,同时我们与中科大附属医院有非常深入的合作,并已经取得非常有临床价值的突破性进展。”
希望有志者加入,共同探索前沿交叉领域,拓展人类知识的边界,为临床医学服务。

参考资料:
1. Duhl, D.M., Vrieling, H., Miller, K.A., Wolff, G.L., and Barsh, G.S. (1994). Neomorphic Agouti mutations in obese yellow mice.Nat Genet 8, 59-65.
2. Manne, J., Argeson, A.C., and Siracusa, L.D. (1995). Mechanisms for the pleiotropic effects of the Agouti gene.Proc Natl Acad Sci U S A 92, 4721-4724.
3. Ollmann, M.M., Wilson, B.D., Yang, Y.K., Kerns, J.A., Chen, Y., Gantz, I., and Barsh, G.S. (1997). Antagonism of central melanocortin receptors in vitro and in vivo by Agouti-related protein. Science278, 135-138.
4. Hahn, T.M., Breininger, J.F., Baskin, D.G., and Schwartz, M.W. (1998). Coexpression of Agrp and NPY in fasting-activated hypothalamic neurons. Nat Neurosci1, 271-272.
5. Cowley, M.A., Smart, J.L., Rubinstein, M., Cerdan, M.G., Diano, S., Horvath, T.L., Cone, R.D., and Low, M.J. (2001). Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus.Nature411, 480-484.
6. Clark, J.T., Kalra, P.S., Crowley, W.R., and Kalra, S.P. (1984). Neuropeptide Y and human pancreatic polypeptide stimulate feeding behaviorin rats. Endocrinology 115, 427-429.
7. Stanley, B.G., Kyrkouli, S.E., Lampert, S., and Leibowitz, S.F. (1986). Neuropeptide Y chronically injected into the hypothalamus: a powerful neurochemical inducer of hyperphagia and obesity. Peptides 7, 1189-1192.
8. Marsh, D.J., Miura, G.I., Yagaloff, K.A., Schwartz, M.W., Barsh, G.S., and Palmiter, R.D. (1999). Effects of neuropeptide Y deficiency on hypothalamic Agouti-related protein expression and responsiveness to melanocortin analogues.Brain Res 848, 66-77.
9. Qian, S., Chen, H., Weingarth, D., Trumbauer, M.E., Novi, D.E., Guan, X., Yu, H., Shen, Z., Feng, Y., Frazier, E., et al. (2002). Neither Agouti-related protein nor neuropeptide Y is critically required for the regulation of energy homeostasis in mice. Mol Cell Biol22, 5027-5035.
10. Gropp, E., Shanabrough, M., Borok, E., Xu, A.W., Janoschek, R., Buch, T., Plum, L., Balthasar, N., Hampel, B., Waisman, A., et al. (2005). Agouti-related peptide-expressing neurons are mandatory for feeding.Nat Neurosci8, 1289-1291.
11. Luquet, S., Perez, F.A., Hnasko, T.S., and Palmiter, R.D. (2005). NPY/AgRP neurons are essential for feeding in adult mice but can be ablated in neonates.Science 310, 683-685.
12. Luo, L. (2020). Principles of neurobiology, (Garland Science).
13. Aponte, Y., Atasoy, D., and Sternson, S.M. (2011). AGRP neurons are sufficient to orchestrate feeding behavior rapidly and without training. Nat Neurosci14, 351-355.
14. Krashes, M.J., Koda, S., Ye, C., Rogan, S.C., Adams, A.C., Cusher, D.S., Maratos-Flier, E., Roth, B.L., and Lowell, B.B. (2011). Rapid, reversible activation of AgRP neurons drives feeding behavior in mice. J Clin Invest121, 1424-1428.
15. Zhu, C., Jiang, Z., Xu, Y., Cai, Z.L., Jiang, Q., Xu, Y., Xue, M., Arenkiel, B.R., Wu, Q., Shu, G., et al. (2020). Profound and redundant functions of arcuate neurons in obesity development.Nat Metab2, 763-774.
16. Chen, Y., Lin, Y.C., Kuo, T.W., and Knight, Z.A. (2015). Sensory detection of food rapidly modulates arcuate feeding circuits.Cell160, 829-841.
17. Mandelblat-Cerf, Y., Ramesh, R.N., Burgess, C.R., Patella, P., Yang, Z., Lowell, B.B., and Andermann, M.L. (2015). Arcuate hypothalamic AgRP and putative POMC neurons show opposite changes in spiking across multiple timescales.Elife 4.
18. Betley, J.N., Xu, S., Cao, Z.F.H., Gong, R., Magnus, C.J., Yu, Y., and Sternson, S.M. (2015). Neurons for hunger and thirst transmit a negative-valence teaching signal. Nature521, 180-185.
运营/排版:何晨龙